|
Pure Genes, Pure Genius
Steven L. McKnight
FullText: http://www.sciencedirect.com/science/article/pii/S0092867412010203
As a physical substance, deoxyribonucleic acid (DNA) was first purified more than 140 years ago by Friedrich Miescher. Some 40 years later, Avery, McCarty, and McLeod utilized purified bacterial DNA to demonstrate it as the transforming substance, thus proving the chemical substance of genes to be composed of DNA. A decade later, Watson and Crick resolved the double-helical nature of DNA. The culminating event in the evolution of this science came roughly two decades later in the form of experiments that facilitated the isolation of purified, single genes. Whereas many scientists contributed to this watershed accomplishment, Don Brown and Tom Maniatis stand in unique positions at or near the apex of the list.
Why would one want to purify a single gene, and what could one do with a purified gene in hand? In retrospect, the answers to these simple questions are obvious. What must be remembered, however, is the fact that, when Brown, Maniatis, and others first accomplished the objective of gene isolation and purification, we did not know how to sequence a gene, we did not know how to express a gene, we did not know where regulatory sequences might be located, and we did not know whether the coding regions of genes would be co-linear with that of their mRNA products. As such, it was an inspired adventure for these scientists to purify genes before we even knew quite what to do with them.
The accomplishments of Brown and Maniatis sit on either side of what may arguably be deemed the most transformative event in biological sciences over the past century, the advent of gene cloning. This happened in the mid-1970s as a result of the science of Daniel Nathans, Hamilton Smith, Tom Kelly, Werner Arber, Herbert Boyer, Stanley Cohen, Paul Berg, and Dale Kaiser. These and other icons arranged the tool kit and provided the assembly directions, facilitating the molecular cloning of single, purified segments of DNA. In the decade before, Donald Brown began to show us what could be done with purified genes. And in the decade after that, Maniatis refined the techniques of gene cloning such that even the least skilled among us could exploit the technology.
Don Brown’s Magic
For historical purposes, I will start with a review of Don Brown’s magic. How could one possibly purify a gene before molecular cloning? Knowing that the density of DNA is proportional to its guanine and cytosine (GC) content, Brown used cesium chloride equilibrium centrifugation to purify the DNA encoding ribosomal RNA. Aside from having a higher GC content than bulk chromosomal DNA, ribosomal genes offered two additional advantages favoring purification. First, the genomes of almost all organisms encode multiple ribosomal RNA genes. Second, as independently demonstrated by Joseph Gall and Brown, ribosomal RNA genes are hugely amplified in number during formation of amphibian oocytes (Brown and Dawid, 1968; Gall, 1968). The combination of these quirky features—high GC content, repetitive nature, and amplification—allowed the independent achievements of Brown and Max Birnstiel to purify ribosomal RNA genes (Brown and Weber, 1968; Birnstiel et al., 1971).
Before DNA sequencing and all of the other capabilities evolving from the field of molecular biology, what could one do with a purified gene? Here is where Brown and others began to show us the way. He began to painstakingly dissect gene anatomy, initially using electron microscopy. By partially denaturing the DNA duplex with a combination of temperature and denaturants, Brown discovered that ribosomal genes are strung along the chromosome in repetitive fashion, with transcribed regions being separated by nontranscribed “spacers.” This discovery was confirmed in spectacular fashion by the elegant electron micrographs taken by Oscar Miller showing actively transcribed ribosomal RNA genes (Miller and Beatty, 1969). Subsequent evolution of this anatomical approach to gene structure employed R looping as a means of showing where on a gene its RNA product belonged and physical mapping via the use of restriction endonuclease enzymes. Although seemingly crude and rudimentary, it was the pioneering work of Brown, Birnstiel, and others that taught us the concept of using physical methods to dissect the anatomy of purified genes.
Brown also exploited the unusually high GC content of the gene encoding silk fibroin of silk moth larvae to purify that gene despite the fact that it was neither repetitive nor amplified. Yet what eventually evolved as Brown’s most rich vein of research came from his purification of the amphibian genes encoding 5S ribosomal RNA. Although repetitive in nature, like the genes encoding the larger 18S and 28S ribosomal RNAs, the 5S ribosomal RNA genes were small and ultimately suited for an unprecedented accomplishment beyond gene anatomy: gene expression. During a decade-long period straddling the advent of gene cloning, Brown and colleagues were able to develop a test tube system that accurately recapitulated 5S ribosomal RNA gene expression (Birkenmeier et al., 1978). This paved the way for the mapping of the regulatory sequences that facilitated 5S gene transcription, which, to the surprise of all, were located right in the middle of each gene (Bogenhagen et al., 1980). Finally, this path of research led to the first discovery of a eukaryotic transcription factor, designated TF3A (Pelham and Brown, 1980). In turn, studies of TF3A led to the discovery of the first zinc finger (Klug and Rhodes, 1987). It can safely be concluded that Don Brown’s path of research laid the groundwork for the entire field of eukaryotic gene regulation, all of which has relied most vitally on purified genes.
Meteoric Rise of Maniatis
With Brown’s pioneering research approaching logarithmic momentum just on the south side of the advent of gene cloning, that of Tom Maniatis was perched just on the north side of the divide. Liberated by cloning, Maniatis did not have to restrict his sights to genes that were repetitive, amplified, or differentiated by unusual GC content—he could study any gene of choice. Despite this liberation, Maniatis employed Brown-like focus in choosing to demystify anything and everything associated with the mammalian genes encoding hemoglobin protein. Maniatis was first to clone a full-length cDNA—that encoding β-globin protein (Efstratiadis et al., 1976) —and first to deduce the structure and sequence of this cDNA (Efstratiadis et al., 1977). These seminal accomplishments laid the groundwork for subsequent studies revealing the identities of human mutations that cause β-thalassemia and methodologically instructed the field as to how to synthesize, clone, sequence, and characterize cDNAs encoding specific gene products. Like Brown, Maniatis taught us how to do it.
The next step in the meteoric rise of “Tom terrific” was his construction of libraries of genomic DNA (Maniatis et al., 1978). Although not alone in accomplishing this feat, few would argue that anyone was more responsible for transforming gene cloning into what we now call genomics. Just the idea that a small test tube could contain recombinant bacteriophage housing each and every segment of DNA in the entire genome of an organism—a virtual library—remains astounding to this day. The value of a library is of course explicitly dependent upon one’s ability to access the individual books or genes. Together with others, Maniatis taught us how to fish out the one-in-a-million recombinant phage that contained our gene of interest—not just the synthetic cDNA copy of a specific mRNA, but also the genomic copy of the very same gene (Fritsch et al., 1980).
It was at this point that the field began to erupt with totally unanticipated surprises. I can vividly remember the very first scientific meeting I ever attended, the 1977 Cold Spring Harbor Symposium on Quantitative Biology. Although nominally organized around the field of chromatin, it was at that meeting where we first learned from Phil Sharp, Rich Roberts, Rich Gelinas, and others that mRNA sequences were not co-linear with their encoding genes, the key observation that led to the discovery of pre-mRNA splicing. Once the dust settled and the symposium volume was published, Maniatis’ influence was pervasive. The way to resolve gene anatomy was simple: clone the cDNA, clone the genomic copy, sort out the relationship between the two, and the task was done.
Like Brown, Maniatis took a love to gene expression. He worked collaboratively with Richard Axel to develop cotransfection methods to stably move purified, recombinant genes into mammalian cells, showing how these methods could facilitate the expression of any protein of choice (Wigler et al., 1979). Boy did this change the world! It fundamentally created the biotechnology industry, allowing Genentech, Amgen, and other companies to industrialize the production of recombinant proteins as therapeutic medicines. Not only did Maniatis facilitate this transformation, but he, like Brown, dug deep into the processes responsible for regulating gene expression. He studied regulatory DNA sequences, including promoters and enhancers. He discovered transcription factors that bound to these sequences. He studied pre-mRNA splicing—anything and everything to do with the enabling technology of having purified genes in hand.
Molecular Recipes that Work
In addition to their profound records of scientific accomplishment, both Brown and Maniatis selflessly contributed to the broad community of biomedical sciences in special ways. Starting with its first edition in 1982, Maniatis teamed up with Ed Fritsch and Joe Sambrook to assemble “Molecular Cloning: A Laboratory Manual” (Maniatis et al., 1982). This and subsequent editions constituted comprehensive cookbooks for the deployment of all methodologies essential for even the most novice of scientists to productively use the techniques of molecular biology. Although it is a testament to the science of molecular biology itself that it can be utilized in so facile a manner worldwide, there is no doubt that the Maniatis, Fritsch, and Sambrook cloning manual liberally greased the wheels for the dissemination of the technology. Just like the cookbooks of Betty Crocker and Fannie Farmer, the molecular cloning manual was chock full of recipes that worked. It was endowed with easily deciphered indices and was likewise updated through the years to incorporate the ever-expanding and improving technologies of the field. It can be argued that no other printed treatise has had more value and impact to biomedical sciences than the molecular cloning manual produced by Maniatis and colleagues.
Creating Something from Nothing
Don Brown’s extracurricular contributions to science come in a very different flavor. Back in the early 1980s Brown founded the Life Science Research Foundation (LSRF). With not a dime in his pocket, Brown decided to create the LSRF as a postdoctoral fellowship foundation. His concept was simple: gather a bunch of friends to offer gravitas (Don Seldin, Jim Watson, Bruce Alberts, Sol Snyder, Phil Sharp, Paul Berg, David Baltimore, Joe Goldstein, Mike Brown, Al Gilman, Shirley Tilghman, etc.), assemble a review team composed of the very best scientists (long headed by Tom Silhavy and Jim Broach), and then badger the profit-making pharmaceutical and biotechnology companies to donate the funds necessary to provide fellowship stipends to 10–20 young biologists every year. The LSRF has now been in operation for a quarter of a century and has funded the postdoctoral training of more than 400 young scientists. From 500–1,000 applications submitted each year, the advisory committee selects LSRF finalists solely on merit—no politics at all. It was Brown’s vision to have the LSRF burdened with almost no restrictions with respect to the citizenship of the applicants and no restrictions as to whether the award would be for a first or second postdoctoral fellowship; the applicants could also work in literally any field of biology or medicine. The legacy of this selfless act of community service is the training and allegiance of hundreds of young scientists—each and every one to whom Don Brown is godfather.
Personal Reflections
Having known and admired Don Brown and Tom Maniatis for more than 30 years, I close with several personal reflections. Both Brown and Maniatis are serious and ambitious men. Their seriousness, however, never succeeded in masking their open and vibrant personalities. Both are people who, upon entry, light up the room. Likewise, their ambition never masked their humanity. Both Brown and Maniatis contributed to the scientific community in ways that matched or exceeded their personal achievements as scientists. Every institution where Maniatis has been employed—from Cold Spring Harbor, to California Institute of Technology, to Harvard, to Columbia—has gained more from Tom than he has from them. It was Maniatis who founded the molecular cloning course at Cold Spring Harbor Laboratory, a course that I had the privilege of teaching and a course that has helped scientists young and old to learn the new techniques of molecular biology. Maniatis did this selflessly—this is Tom Maniatis.
It can likewise be said that the fingerprints left by Don Brown on the Department of Embryology of the Carnegie Institution of Washington remain indelible. From the early 1970s to near the end of the century, Brown created and directed the institutional paradigm of using pure genes to pry apart complex biology. Along with Gerry Rubin, Allan Spradling, Joe Gall, Andy Fire, Nina Fedoroff, Alejandro Sanchez-Alvarado, Doug Koshland, and many others, I was one to profit from Don Brown’s inspirational leadership. Offering one small personal reflection, Don let me work as a nominally “independent” postdoctoral fellow. During a 3 year period, I endeavored to pick apart the promoter of the herpes simplex virus thymidine kinase (TK) gene via a process that I dubbed “linker scanning mutagenesis.” Brown coached me all the way, but when it came time to publish the work (McKnight and Kingsbury, 1982), Don refused to be listed as an author. That single act of generosity affected me more profoundly than all of the technical experience garnered during my postdoctoral training.
Through their science, Don Brown and Tom Maniatis taught us how to use and deploy pure genes. Through their commitment to the scientific community, Don and Tom taught us the pure genius of giving back. It is hard to conceive of a better duo to receive the 2012 Albert Lasker Special Achievement Award in Medical Science.
Acknowledgments
I thank Drs. Joseph Goldstein and Joseph Gall for editorial comments helpful to the revision of this essay.
References
E.H. Birkenmeier, D.D. Brown, E. Jordan Cell, 15 (1978), pp. 1077–1086
M.L. Birnstiel, M. Chipchase, J. Speirs Prog. Nucleic Acid Res. Mol. Biol., 11 (1971), pp. 351–389
D.F. Bogenhagen, S. Sakonju, D.D. Brown Cell, 19 (1980), pp. 27–35
D.D. Brown, I.B. Dawid Science, 160 (1968), pp. 272–280
D.D. Brown, C.S. Weber J. Mol. Biol., 34 (1968), pp. 681–697
A. Efstratiadis, F.C. Kafatos, A.M. Maxam, T. Maniatis Cell, 7 (1976), pp. 279–288
A. Efstratiadis, F.C. Kafatos, T. Maniatis Cell, 10 (1977), pp. 571–585
E.F. Fritsch, R.M. Lawn, T. Maniatis Cell, 19 (1980), pp. 959–972
J.G. Gall Proc. Natl. Acad. Sci. USA, 60 (1968), pp. 553–560
A. Klug, D. Rhodes Cold Spring Harb. Symp. Quant. Biol., 52 (1987), pp. 473–482
T. Maniatis, R.C. Hardison, E. Lacy, J. Lauer, C. O’Connell, D. Quon, G.K. Sim, A. Efstratiadis Cell, 15 (1978), pp. 687–701
T. Maniatis, E.F. Fritsch, J. Sambrook Molecular Cloning: A Laboratory Manual Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (1982)
S.L. McKnight, R. Kingsbury Science, 217 (1982), pp. 316–324
O.L. Miller Jr., B.R. Beatty Science, 164 (1969), pp. 955–957
H.R.B. Pelham, D.D. Brown Proc. Natl. Acad. Sci. USA, 77 (1980), pp. 4170–4174
M. Wigler, R. Sweet, G.K. Sim, B. Wold, A. Pellicer, E. Lacy, T. Maniatis, S. Silverstein, R. Axel Cell, 16 (1979), pp. 777–785
On Getting There from Here
Steven L. McKnight
FullText: http://www.sciencemag.org/cgi/content/full/330/6009/1338?ijkey=06Az947.SW9ZE&keytype=ref&siteid=sci
Studies on a variety of interesting biological problems, ranging from circadian rhythm to cancer cell growth to longevity, have begun to give evidence that the physiological state of cells and tissues reflects both the cell’s regulatory systems and its state of intermediary metabolism. It is appreciated that the regulatory state of a cell or tissue, as driven by transcription factors and signaling pathways, can impose itself upon the dynamics of metabolic state. It follows that the reciprocal must also be the case, that metabolic state will feed back to impose itself on regulatory
state. An appreciation and understanding of this reciprocity may be required to crack open problems in biological research that have heretofore been insoluble.
For the past 30 years, research in the biological sciences has been dominated by molecular biology. The successes of this approach have shaped our understanding of innumerable domains of biology. But any field that becomes sufficiently muscular can overshadow other credible approaches to scientific inquiry. One field etiolated by the cloud of molecular biology has been metabolism. The vast majority of discoveries made by molecular biologists over the past several decades required no attention to the metabolic state of a cell. Molecular biologists needed no distracting
thoughts about the metabolic state of a cell to discover microRNAs, the reprogramming of somatic cells into pluripotent stem cells, or gene
rearrangement as the underlying basis for the generation of antibody diversity.
One simple way of looking at things is to consider that 9 questions out of 10 could be solved without thinking about metabolism at all, but the 10th question is simply intractable. As the saying goes, you simply “can’t get there from here” to answer this 10th question if you are ignorant about the dynamics of metabolism. This, I propose, is where we are now finding pregnant opportunities in the field of experimental biology. The low-lying fruit that could be picked by molecular biologists without having to consider the metabolic state of a cell, tissue, or organism is largely gone. The more sticky problems that required attention to the dynamics of metabolism and that were pushed aside for decades now loom as interesting and important challenges.
Consider a prime example of how molecular biologists have begun to embrace the importance of metabolic regulation. Cancer researchers have long known of the enigmatic ability of tumor cells to undertake aerobic glycolysis, the so called Warburg effect (1, 2). It makes sense that cancer cells would be highly glycolytic, yet why would these cells choose to dispose of the terminal product of glycolysis? Instead of allowing pyruvate to be converted into acetyl–coenzyme A (CoA) via the spectacularly beautiful pyruvate dehydrogenase enzyme complex within mitochondria, cancer cells instead convert pyruvate into lactate through the lactate dehydrogenase enzyme, and then simply excrete it—blindly giving away exceptional energetic value stored in the lactate hydrocarbon. Any cell that wants to grow—and there is nothing cancer cells care more about than growth—would be crazy to waste hydrocarbon; this would be akin to a motorist driving down the New Jersey Turnpike throwing away gasoline. The spendthrift waste of lactate likewise deprives the cancer cell of huge amounts of acetyl-CoA to be used for the synthesis of lipids, sterols, and other cellular building blocks. Despite progress, attention, and plenty of hype, it is safe to conclude that the famous Warburg effect remains a mystery.
Cancer researchers now recognize that regulatory proteins, such as the hypoxia-inducible transcription factors, can directly regulate the expression of genes encoding glycolytic enzymes (3). They now pay attention to how their favorite regulatory proteins, including the Myc and p53 transcription factors, help set the metabolic state of cells. The fact that thesemasterful transcription factors participate in dictating the metabolic state of a cell is now beginning to be accepted. The equally compelling corollary, however, remains largely unappreciated. That is, if regulatory state (transcription factors, signaling pathways, etc.) is accepted to control metabolic state, is it not also unconditionally certain that metabolic state will reciprocally control the regulatory state itself? Understanding this reciprocity, and digging to the bottom of it, is where the future lies. Perhaps fittingly, this research will require the sophistication of scientists having genuine skills in the study of enzymology and intermediary metabolism.
Whole-genome sequencing efforts of individual tumors, now numbering in the thousands— yet soon to be numbered in the hundreds of thousands—are providing unbiased views of the myriad of “oncogenotypes” that underlie human cancer. Instead of ignoring mutations that happen
to fall in the genes encoding metabolic enzymes, scientists seem more keen than ever to find and study such mutations. This change may reflect
the recognition by cancer researchers that mutations that alter the function of metabolic enzymes might help to resolve the enigmatic, aerobic glycolytic state of certain cancer cells. It is equally likely that an understanding of how cancer cells veer away from normality with respect to intermediary metabolism might lead to the conceptualization of new and inventive strategies for therapeutic intervention.
For example, a set of recurrent genetic lesions believed to influence the metabolic state of glioblastoma cancer cells have been identified in the
genes encoding either of the two isoforms of isocitrate dehydrogenase (4, 5). Perplexingly, most, if not all, of these lesions mutate a single arginine
residue in either of the two isoforms of the enzyme (IDH1 or IDH2). The precise selectivity of the mutational events, coupled with the observation that only one allele of either enzyme appears to be mutated in human cancer, pointed to the possibility that the lesions might be causing the enzymes to adopt a new catalytic function. Indeed, the mutated forms of the IDH1 and IDH2 enzymes exhibit a reduced affinity for isocitrate and are endowed with a new catalytic functionwherein a-ketoglutarate is converted in an NADPH (nicotinamide adenine dinucleotide phosphate, reduced)– dependent manner to 2-hydroxyglutarate (6).
What pathways might be expected to be altered in a cell impeded for the production of aketoglutarate and concomitantly endowed with
increased intracellular production of 2-hydroxyglutarate? A logical guess would be the family of dioxygenase enzymes that use a-ketoglutarate as
an essential cofactor and, simultaneously, are inhibited in the presence of 2-hydroxyglutarate. Included among this family of dioxygenase enzymes
are the prolyl-hydroxylase enzymes that modify and negatively regulate the HIF-1a (hypoxiainducible factor 1a) andHIF-2a transcription factors
in oxygenated cells (7, 8). Partial elimination of these dioxygenase enzymes could be interpreted to lead to the activation of the hypoxia response
pathway, thereby accounting for the activation of transcription of genes encoding glycolytic enzymes just as happens in the absence of the Von
Hippel–Lindau (VHL) tumor suppressor gene (9, 10). This interpretation is very likely oversimplistic. Attenuation of a-ketoglutarate concentrations
and accumulation of increased amounts of 2-hydroxyglutarate almost certainly lead to the inhibition of other dioxygenase enzymes, of which mammalian cells have scores of isoforms. Intriguingly, some of these additional dioxygenase enzymes have been implicated in the control of histone demethylation (11–13), leaving open the possibility that changes in metabolic state might impose alterations in the epigenetic state of cancer cells.
The drift of this thinking is concordant with the recent discovery of mutations found in renal cell cancers in the mitochondrial enzyme fumarate
hydratase that converts fumarate to malate (14). These mutations are more rare and of a recessive nature—wherein both alleles of the gene
encoding fumarate hydratasemust be inactivated. Because fumarate is known to be an inhibitor of the aforementioned family of dioxygenase enzymes, it is conceptually logical to hypothesize that the accumulation of excessive amounts of fumarate might inactivate the prolyl-hydroxylase
enzymes that normally keep the HIF transcription factors in an inactive state (and, perhaps, likewise impose alterations in the epigenetic state of cancer cells). Finally, the same reasoningmight apply to rare mutations in the human genes encoding mitochondrial succinate dehydrogenase and its
assembly factors that are believed to contribute to the formation of paragangliomas and pheochromocytomas (15, 16). The exciting angle on these
studies of cancer-causing mutations in the genes encoding isocitrate dehydrogenase, fumarate hydratase, and succinate dehydrogenase is that they
formally predict the concept that the metabolic state of a cell can indeed exert control over its regulatory state, thereby confirming the reciprocal relationship between the two.
One enduring complication that shows little sign of resolution concerns the manner in which cancer cells are grown and studied in tissue culture
plates. We routinely grow cancer cells under conditions of unlimited access to glucose—not to mention oxygen, vitamins, known and unknown growth factors present in serum, and every nutritional fertilizer imaginable. The growth environment of cancer cells within tumors, especially solid tumors, could hardly be more different than that of cells being grown under standard, tissue culture conditions. It is reasonable to suspect that cancer cells weave their way through exceptionally selective oncogenetic gymnastics to achieve a growth-permissive metabolic state. If so, what features of this metabolic state can be anticipated to be preserved and studied when cells are practically grown in Karo syrup or alongside floating logs of Snickers Bar candy? Returning to the “you can’t get there from here” theme, it is predictable that we will have to find more biologically sound ways to grow cancer cells in culture in order to favorably use them in efforts to discover therapeutics that might exploit their unique metabolic state.
The resurrection of research involving or including metabolism is clearly upon us—that is the good news. The bad news is that the field was sufficiently snuffed over the past several decades that we have precious few scientists who have been trained to genuinely understand intermediary metabolism. Just because we can now pronounce the names of the metabolic enzymes whose encoding genes and mRNAs show up on
our ChIP-Seq (chromatin immunoprecipitation– sequencing) and DNA microarrays lists does not necessarily mean that we can put two and two
together. Despite the handicap of not being able to field an experienced team at this point, it is encouraging to see favorable trends that may enable negotiation of discovery routes that were, until now, largely obscure.
References and Notes
1. O. Warburg, K. Posener, E. Negelein, Biochem. Z. 152, 319 (1924).
2. O. Warburg, Science 123, 309 (1956).
3. N. V. Iyer et al., Genes Dev. 12, 149 (1998).
4. D. W. Parsons et al., Science 321, 1807 (2008).
5. H. Yan et al., N. Engl. J. Med. 360, 765 (2009).
6. L. Dang et al., Nature 462, 739 (2009).
7. A. C. R. Epstein et al., Cell 107, 43 (2001).
8. R. K. Bruick, S. L. McKnight, Science 294, 1337 (2001).
9. P. H. Maxwell et al., Nature 399, 271 (1999).
10. M. Ohh et al., Nat. Cell Biol. 2, 423 (2000).
11. Y. Tsukada et al., Nature 439, 811 (2006).
12. J. R. Whetstine et al., Cell 125, 467 (2006).
13. P. A. Cloos et al., Nature 442, 307 (2006).
14. J. S. Isaacs et al., Cancer Cell 8, 143 (2005).
15. M. A. Selak et al., Cancer Cell 7, 77 (2005).
16. H. X. Hao et al., Science 325, 1139 (2009).
17. I thank M. Brown, R. Bruick, B. Tu, and J. Rutter for helpful editorial comments.
Unconventional Wisdom
Steven L. McKnight
DOI 10.1016/j.cell.2009.08.016 (Download pdf)
"May your hands always be busy,
May your feet always be swift,
May you have a strong foundation
When the winds of changes shift.
May your heart always be joyful,
And may your song always be sung,
May you stay forever young"
Bob Dylan - 1973
When asked to compose a brief article to be directed toward an audience consisting of young scientists, my first reaction was—geez, young scientists don’t need advice at all, they rule. The majority of genuinely profound discoveries made over the millennia have come from young scientists. There are numerous reasons for this. The young mind is maximally acute, and the young scientist is minimally distracted. It’s sink-or-swim for the young scientist, so he or she is hungrier and fights harder to bring home the bacon. Finally, the young scientist brings little baggage to a problem. I would trade the supposed wisdom gained from a longstanding career in science for the combination of naiveté and exuberance any day. For these reasons, the best advice I can give comes from one of Bob Dylan’s songs on his Planet Waves album: “may you stay, forever young.” The starting point I was given posed the question of where I thought the best opportunities for discoveries might lie in the years ahead. The answer to this question is simple—scientific opportunities abound everywhere. Of course, we hope to someday understand the molecular basis of memory and the magic of regeneration, but these are only two of hundreds of watershed opportunities for transformative discovery standing in front of us in the decades to come.
Let’s consider two contrasting articles, published by eminent scientists, that influenced me early during my own career. The first was authored by the renowned geneticist Gunther Stent. Having used Stent’s wonderful textbook on genetics (Stent, 1963) during my undergraduate training at the University of Texas, I viewed Stent as a scientific icon. Subsequently, Stent published an essay in Science that argued that the field of molecular biology was washed up, done, kaput (Stent, 1968). Yes, by then the genetic code had been cracked, Crick’s central dogma had been confirmed, and the nature of the gene was understood in atomic detail. On the other hand, I was just entering the field of biological research, and I viewed the subdiscipline of molecular biology as having unlimited promise. Much to my dismay, a scientist of Stent’s stature had argued the field was dead.
The second article of influence was an essay published in Cell by Charles Yanofsky wherein he questioned whether the field of microbiology might be at the end of its rope (Yanofsky, 1991). At the time, the lion’s share of attention and grant funding were going toward studies of eukaryotic organisms, and Yanofsky wondered whether the end was near for studies of prokaryotes. Yanofsky, however, came to the exact opposite conclusion
from Stent—instead of announcing the demise of microbiology, he boldly predicted that the well was far from dry. Looking back on what has been discovered in the field of microbiology over the past three decades—quorum sensing, molecular mechanisms of pathogenesis, riboswitches, genome sequences, thermophiles and extremophiles, the microbial flora inhabiting our bodies, and so on—Yanofsky’s forward-reaching conclusion could not have been more spot on. Naming where the most exciting breakthroughs will come from in the ensuing decades is way beyond what I could possibly muster! In this regard, I pay special homage to an early president of the Carnegie Institution of Washington, Vannevar Bush. It was his words that so beautifully described science as the “endless frontier” (Bush, 1945). Those two words gave me plenty of inspiration to overcome any discouragement transiently resulting from the diametrically opposing conclusion of Stent. They are as true today as they were in 1945 when penned by Dr. Bush.
Reasoned advice to the young scientist is to be careful not to become overly focused. Yes, to be competitive, the young scientist must be at the top of the game in his or her chosen field. On the other hand a scientist broadly exposed to disciplines outside of his or her chosen field will enjoy distinct advantages. The subdisciplines of biological and biomedical research evolve rapidly, and it is often the case that the most radical of transformations to a field come from outsiders who bring a combination of fresh perspectives and naiveté. It is for this reason that medical students—if they choose in a genuine and dedicated manner to have a career in science— can sometimes be equally prepared for extended success as PhDs. Medical students have to learn anatomy, physiology, pathology, genetics, biochemistry, pharmacology, immunology, and other fields of science required to understand how the human body operates. Modern PhD programs often focus the training of young scientists so acutely that, as a subdiscipline matures, liability to extinction becomes a genuine threat. For the PhD student, one should consider the benefits of attending seminars—while in graduate school and during postdoctoral training—orthogonal to what is being studied in the training laboratory. The much stronger tendency for a young trainee is never to miss the seminars most closely related to his or her research, even though the young scientist already has a 99% mastery of that particular subdiscipline. Bottom line, the breadth of your scientific training will be of equivalent value to its depth.
The question is not where to explore for new opportunities on the horizon of science, but instead, how to go about looking for them. Here the balance of “inductive inquiry” (I2) and “hypothesisdriven” (HD) approaches becomes the crux. A recent article written by Francisco Ayala beautifully recounts how the plusses and minuses of the two approaches have been debated by philosophers over the past several centuries (Ayala, 2009). The I2 approach, in its most pure form, entails adventure into uncharted territories—neither guided nor bridled by hypothesis. The HD approach is built on scholarship and smarts and is fundamentally driven by theory. If X and Y facts are understood, this knowledge should facilitate the hypothesis essential for solving the unknown Z. Scientists, just like most people, are far more comfortable with the known than the unknown. If one can embark on an adventure with known variables in pocket, the comfort factor alone will nudge the endeavor in favor of the HD direction.
Another influential factor perennially favoring the HD side of the equation is money. Except in the most unusual of circumstances, other people are making the decision for us as to where we get to explore. Science requires money, and money is doled out by committees that evaluate our research plans. Proposals thin on HD, no matter how open and uncharted the territory chosen for inquiry, tend to be rejected. As such the I2 approach almost always loses out to the HD approach when it comes to funding decisions. Bottom line, conventional wisdom almost always prevails—this, I advise, is something you will have to constantly fight in order to carve out a truly innovative career in science.
Any research endeavor we might choose to pursue is, of course, an I2/HD blend. As articulated by Charles Darwin, “Let theory guide your observations,” otherwise one “might as well go into a gravel pit and count the pebbles and describe the colors. How odd it is that anyone should not see that observation must be for or against some view if it is to be of any service” (Darwin, 1903). A scientist cannot easily shed the knowledge causing him or her to proceed on an adventure without bearing elements of scholarship, theory, and bias. If, however, this knowledge is sufficiently acute and dominating, it likely leads the adventure in the same direction being pursued by many other scientists. Put in other words, the dominating hypotheses in all fields—like the Pied Piper of Hamlin—tend to channel scientists into the same directions. A buffering of the power of the HD approach requires a purposeful squinting of the eyes so that a dose of I2 flavor can be added to the mix. Please understand that I am not advocating the mindless data gathering that has become trendy with the advent of “omics” technologies (DNA microarrays, whole genome association scans, and the like). These approaches do little more than count and color Darwin’s pebbles. What I instead recommend is fresh scientific inquiry into under-appreciated biological or medical phenomena that presently exist in a mystic state.
Central to my argument favoring inductive inquiry is the attitude that we know so little about biology that we cannot even anticipate the nature of major discoveries to unfold in the future. Others, no doubt, are more perceptive than I. But I can legitimately say that I had no clue that eukaryotic genes would be segmented into introns and exons, that RNA could perform catalytic reactions, and that small RNAs would be able to self-amplify and profoundly regulate biological pathways in organisms ranging from spinach to worms to humans. Cast in a different light, I pose the question of whether our biomedical research enterprise would be better or worse off had every single specific aim of every single grant ever submitted to the NIH been perfectly completed if—in payment to the devil—we had to give up the totally unanticipated discoveries that were never once written as a specific aim in any grant application?
So, if one buys into the utility of the foggy, eye-squinting I2 approach, how might a young scientist pursue this course and decide what to do? The actual choice of direction is the easiest problem to solve. One simply has to look where the trends are headed and go the other way. Here, for conceptual purposes only, I suggest a “pin the tail on the donkey” approach. A randomly assembled chart is printed up containing squares labeled with all of our 20 to 30 thousand genes. Of these, we know lots about some, a bit about others, yet almost nothing of the remainder. We slap on a blindfold then throw the dart against the wall. Chances are reasonably good that the dart will land on an “unknown” gene—as long as the contestant does not peek around the blindfold and aim the dart at the squares adorned with the comfortable names that already appear every day in the literature. That the unknown gene does something critical is supported by the fact that it’s been kept in place by hundreds of millions of years of evolution. That every gene and every protein are both interesting and important is incontrovertible. This being the case, why would anyone want to work on a gene or protein already staked out by dozens of other scientists?
Why do we choose to be scientists? Most fundamentally, we do so because science offers us the chance to make a discovery—no matter how large or small—never before conceived by another human. Two hundred years ago, the opportunity for discovery is what drove a band of adventurous souls to join Meriwether Lewis and William Clark to sail up the Missouri river in hopes of finding a passage across the northwest. Nothing, I propose, can be more rewarding than the sheer joy of discovery. It is notable, however, that those mavericks who signed on with Lewis and Clark experienced 99% slog to the 1% of their time spent miraculously stumbling over new valleys or passages. Scientific research, likewise, is a head-bumping slog. If we are lucky, the slog is periodically punctuated by unbridled joy. In this time of tight grant funding and a challenging job market, the best I can offer is to encourage young scientists to trust your instincts and stay on your uniquely chosen path.
I close with a personal reflection. When I was a youngster, I loved sports and could think of nothing better than a career in professional athletics. The reason for this was not based on talent— had it been, I’d perhaps now be a retired football player coaching at some high school or college. No, the reason for this was that I simply loved sports. I was unafraid of training and working to foster my ambitions for achievement because it never felt like work at all. As, through adolescence, I came to realize that my innate talents in athletics were clearly inadequate for a professional career, I was haunted by the question of what I might do for a living. Out of serendipity, I found my way into the field of biological research. Lo and behold, I found that chasing scientific adventure was hardly work at all but instead was a joyous endeavor not unlike what I’d experienced at an earlier stage of my life in athletics. To the young scientist, I leave this final question. Does science feel like a job, or is it the case that vocation matches avocation, such that you can’t wait to get to the lab, such that it does not feel like work at all? If so, nothing can stop you and may you indeed “stay, forever young.”
Acknowledgments
I thank Cori Bargman, Don Brown, Joanne Chory, Mike Dyer, Charlie Emerson, Joe Goldstein, Rich Losick, Mort Meyerson, Mark Ptashne, Bill Neaves, Mike Rosen, Peter Walter, Xiaodong Wang, Charley Yanofsky, and trainees in the McKnight lab for invaluable input on the composition of this Essay.
References
Ayala, F.J. (2009). Proc. Natl. Acad. Sci. USA 106, 10033–10039.
Bush, V. (1945). Science, The Endless Frontier: A Report to the President by Vannevar Bush, Director of the Office of Scientific Research and Development (Washington, DC: United States Government Printing Office).
Darwin, F. (1903). More Letters of Charles Darwin (London: Murray).
Stent, G.S. (1963). Molecular Biology of Bacterial Viruses (San Francisco: W.H. Freeman and Co.).
Stent, G.S. (1968). Science 160, 390–395.
Yanofsky, C. (1991). Cell 65, 199–200. |
King of Blue
Steven L. McKnight - August 2009
Unexpected things happen in life, that’s about all I’ve learned. Some events are positive – others are negative. I’m about to see my 60th birthday as I write this story. It’s the autumn of my life and as good a time as any to look back.
Like most people, I prefer to look back to the good rather than the bad – but the good is always embedded in the bad and vice-versa. It’s also the case that stories can’t be told in a vacuum, so I start this story with my processing “in country” at the 90th replacement center in Long Binh, Vietnam. I’d performed poorly in my first year of college. 1968 was a bad year to flunk out of college. I was outfitted with flack jacket, helmet, poncho liner and M16 rifle in Xuan Loc, assigned to a tank battery at Phu Loi, and deposited into my assigned tank section in Phuc Vinh – in proximity to a region called the Iron Triangle.
My fatigues were bright green, not having had time to fade into the colors of the uniforms worn by the old-timers who knew the score. My boots were shiny, no time to wear into scruff. My skin was pale and soft - all beacons defining the looks of an FNG – fucking new guy. Like FNG’s before me and after, I was avoided with disdain by everyone except other FNG’s.
Two tanks per section, five guys per tank – commander, driver, gunner, two ammo loaders. Tanks were 26-ton relics from world war two, called dusters - two 40 millimeter pom-pom guns protruding from a washtub-like turret. Everyone but the driver sat on top – inside was a steel coffin where death came from land mines or rockets. Tanks were worthless. The Viet Cong (VC) in the Iron Triangle had long since figured them out. Whether protecting, ostensibly, a convoy – or accompanying a platoon of infantry in random jaunts through the jungle – tanks made two things, a bunch of racket and a ripe target. The VC could hear tanks from miles away. They knew how to hide, which is easy in the jungle, or simply blow the tank up with a land mine or a rocket propelled grenade. An incessant air campaign gave the VC a steady supply of ordinance for land mines in the form of bombs that failed to explode. The Russians and Chinese gave the enemy plenty of rocket propelled grenades. Tanks were worthless.
I didn’t know these things – or much of anything else pertinent to survival. Old timers did. Lots to learn, 99% being on-the-job. Take your daily prophylactics for malaria and have a steady case of the runs. Skip them, get mosquito bit, get the fever, and get choppered to the 93rd evacuation hospital. Sticking with acumen for the shortcut, I chose and enjoyed/suffered the latter option.
A Cajun from Lousiana commanded the tank, a strapping guy from rural Washington was the gunner, a black guy from Georgia was the driver. Jimmy Poole – another Lousiana coon ass – joined me as an ammo loader and FNG partner. Our only other companion was a feral dog – tank mascot – Sam. For army guys, the tour of duty was one year. The Cajun tank commander had been in country nearly that long, just about as hard-bitten, just about as far from FNG status as a draftee could get. The driver and gunner were somewhere in-between – Poole and I had a whole year in front of us.
The feral dog had been with the tank for three or four years. Soul was a better description of Sam than mascot. He’d seen 3 or 4 full rotations through that one tank – seen FNG’s rotating through the cycle over and over and over. Sam and the tank were the constants, the rest of us just transients. Sam’s master was Jordan, the easy-going tank driver. Sam, unlike the old timers, showed me no disdain. He’d look me in the eye and not avoid me. As you’ll see, Sam had magic.
The first six months went fast. My FNG status was resolved a month or so into the tour. The tank crew had to make a run into a jungle fire support base (FSB) being clobbered, had to bring them a transmission for a broken tank that needed fixing before the shit-hole could be evacuated. We flipped a coin, lost the call – meaning our tank had to lead the rescue convoy (the other tank of the section won the coin flip and ran slack at the rear of the convoy). Our destination was FSB Dacus, named after an infantryman killed in a stay-behind ambush earlier in the war. We headed out of Phuc Vinh up toward highway 14 and veered off at Dong Xoai onto a logging road leading into the heart of War Zone D.
Troops at Dacus had spilled the beans to local villagers that the towel was being thrown in – the FSB would be evacuated any day. The VC were connected and resourceful. Anticipating withdrawal, they set up an ambush on the jungle road several miles outside of Dacus. They buried a land mine where a culvert bridge spanned a stream, put bunkers backed up into the jungle on either side of road. First vehicle over the stream gets whacked by the mine, convoy stops - pinned in on either side by the jungle, ambush chews it up.
The luck of it was that the VC didn’t know that one of the tanks at FSB Dacus was busted. Our rescue convoy approached the ambush from the wrong direction. Rumbling down the jungle road, our tank crew saw a stick protruding from a puddle – paper note impaled atop the stick. We got a Kit Carson scout (VC turncoat) to read the note – “beware Vietnamese civilians, do not use this road today” – spooky. We called this up to the colonel flying round and round above us in a command and control helicopter, sage advice – keep going.
A mile or two up the road we came upon the stream and culvert where the VC had buried the mine. Infantrymen walking alongside our tank saw bunkers on both sides of the jungle road just beyond the stream. Get that tank up here and blow those bunkers. We loaded up the pom-pom guns, rumbled over the culvert – boom – we’d sprung the ambush from the wrong direction. People, rifles, ammo, dog flying up in the air. I landed upside down, dazed – ran into jungle. Helicopter gun ships firing, the screaming firefight was over in no time. VC were smart, bolted – they had no purchase on an ambush sprung from the wrong direction. The convoy limped into Dacus that night sans one tank. I was hurting, it rained all night and I slept in a foxhole filled to the brim with water. Next morning we put a new transmission in the broken tank, abandoned Dacus by backtracking out of the jungle. No longer was I an FNG, action/wounds trumped both innocence and limited time in country.
After six months of boredom punctuated, infrequently, by violence – Jordan – Sam’s master, rotated home. Sam chose me to replace Jordan. It’s 1970, American involvement peaking in 1968/69 had begun to decline. Our unit was shut down. We drove the tanks back to Long Bihn and petrified them in cosmoline for use in a future war. Poole, Sam and I got reassigned to a new unit. Irrespective of Nixon’s benevolence in “Vietnamizing” the war, we still had to finish out our tour of duty.
Poole, Sam and I loaded up in the bed of a duce and a half with a ragtag crew, departed Long Binh – headed west through Cu Chi and Go Dau - genuine “Indian country”. We veered away from highway 22 at Tay Ninh (rocket city) then headed west till dead-ending at fire support base Blue, smack on top of the Cambodian border. I’m guessing it was a 6 hour ride with twists and turns, who could have cared – someone else was driving the truck. The mood at Blue was mean, it had been overrun a month earlier by VC sappers – tanks and guns blown up, a bunch of GI’s killed.
The hardscrabble character of Blue was embodied in Mutt, the mascot of the artillery unit occupying the fire base. Never seen a meaner looking dog – adorned with a face scarred by a life of fighting. As we hopped off the truck, Mutt tore into Sam with fury. The local GI’s cheered Mutt on while Poole and I watched helplessly. Lo and behold Sam whipped Mutt. Poole, Sam and I dug a hootch and settled in for the night as unwelcome guests.
Next morning Poole and I were sent out on some worthless recon mission. Upon returning I started hunting up Sam – no luck. Some lifer gunnery sergeant told me that he thought he saw Sam being loaded on a resupply truck heading back to Long Bihn to pick up more ammunition for the big, eight inch guns. I tracked this story up to the captain of the artillery unit – yes, he told me that one dog was enough on FSB Blue – yes, he’d ordered Sam to be shipped out on the resupply run and abandoned along the way. I went flame-throwing mad, told the son-of-a bitch that I was going to kill him – dove at his throat and tried to choke him to death.
Got pulled off and calmed down enough to explain what Sam meant to me and all the guys who’d preceded me back on that rumbling tank blown up earlier in my stay. The artillery captain agreed to let me take off on the resupply run the following morning – drivers showed me where Sam had been kicked off. It was Go Dau – legitimate Indian country. They dropped me off and continued their way back to Long Binh for resupply, I spent the day searching the environs of Go Dau for Sam – lone, stupid GI with his M16, flack jacket and helmet looking for an abandoned dog. No luck by the time the resupply truck picked me on their way back to Blue – three or four hours of zig-zagging our way through jungle roads we rumbled back into Blue. Although I’d reconciled myself into letting the cock sucking captain live, everyone at Blue knew the story and the score. Sam had kicked Mutt’s ass, the captain – while I was on recon – had sealed Sam’s fate. I still had Jimmy Poole as my best buddy, but I’d managed to lose Sam – what a fucking bummer.
Two days later Poole and I were sitting in a bunker with a couple of other GI’s, settling in for the first watch of perimeter guard duty for the night. The bunker happened to face east, back towards Long Bihn. Not believing our eyes, here came Sam trotting right back into FSB Blue. The news spread over the firebase like wildfire – how could that dog, having spent less than a day at Blue – find his way back through 50 kilometers of intersecting jungle roads! No more fights with Mutt, Sam was the accepted “King of Blue”. Other than the birth of my kids, and the time my son made a spectacular interception in the Texas state high school football championship game, no time of my life compares with the sight of Sam trotting up to my bunker at FSB Blue.
The next three months slogged on in an uneventful manner, and it then came time for the artillery unit at Blue to be shut down. Knowing that Poole and I were to be assigned to a machine gun unit in the Central Highlands for the final months of our respective tours, and knowing that Sam could not join us in that setting, I convinced the captain to let me go back to Saigon to find a way to ship Sam back to the United States. Got him the shots and papers, paid two months of wages to Pan Am to fly Sam home, saw him off, and shipped off with Poole on a helicopter taking us up into the Annimite mountains for our final duty. The Army lost track of both Poole and me, so we had no mail for those final three months. The joyous day of departure finally came – boarded a flight out of Cam Ran Bay and made it home safe and sound. First call to family broke the news – Pan Am had let Sam out of his travel cage in San Francisco and lost him.
I’ve been haunted ever since. Would it have been best to leave Sam in Vietnam where everything was unraveling and he had no tank and crew to call his own? Or was it better that he be given the opportunity to find his way in a foreign land unforgiving of his skills. San Francisco airport is a bleak place, but Americans don’t eat dog. I, too, wonder whether the best of me was left in Vietnam – a place where neither ambition nor politics meant anything – a place and time where life was lived close to the bone – a place where one got to meet the likes of the King of Blue.
|
Sam - AKA "King of Blue" - with Jimmy Poole at Fire Support Base Blue - circa 1970 |
Postscript:
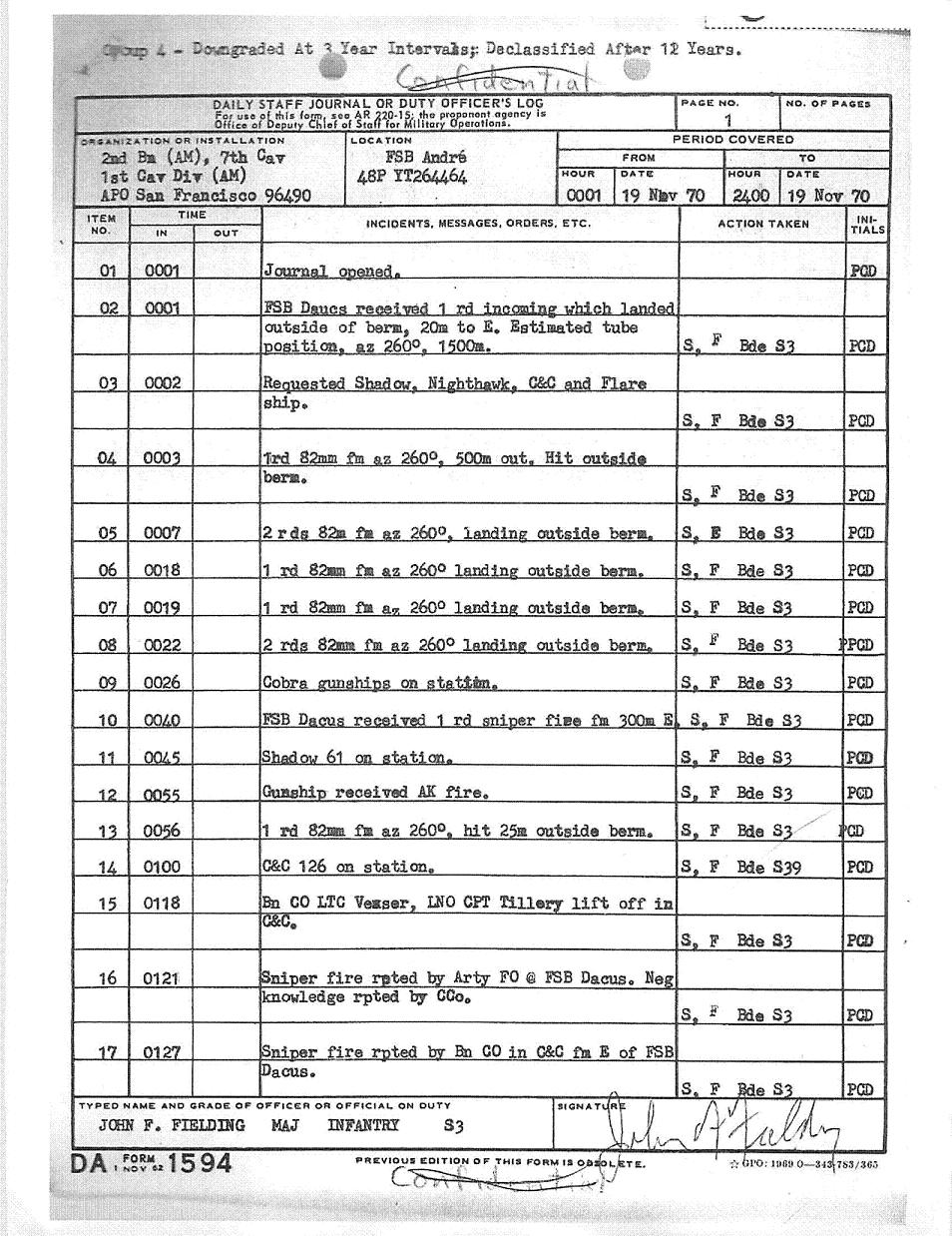
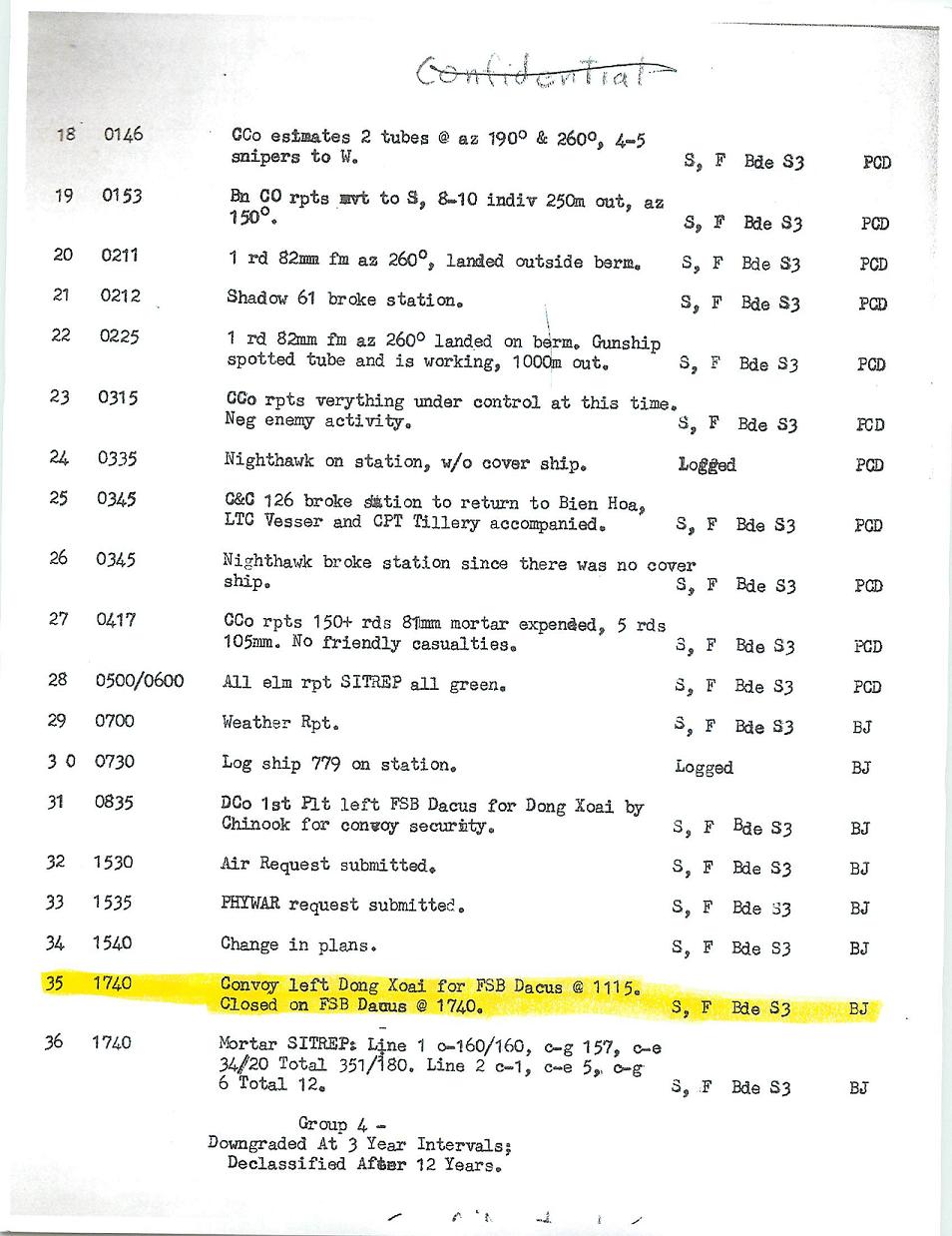
In September of 2011 Dr. McKnight received the following e-mail from Mr. Kevin Perrier. Mr. Perrier, as a member of the First Air Calvary, was at Fire Support Base Dacus at the very time when Dr. McKnight's tank was blown up in the ambush described in "King of Blue". In subsequent e-mail messages Mr. Perrier provided Dr. McKnight with photographs taken at or around FSB Dacus in 1970. Remarkably, one of the photographs actually shows the tracks of the tank that was blown up that very day. Mr. Perrier also provided Dr. McKnight with the "Daily Staff Journal or Duty Officer's Log" from the US Army on November 19th, 1970. Item 35 chronicles the departure of the fated convoy from Dong Xoi to FSB Dacus, which departed Dong Xoi at 11:15AM and arrived at FSB Dacus at 17:40PM.

|